Is the multicellular genome quantum non-local?
A.Einstein and his colleagues, B.Podolsky and N.Rosen [Einstein, Podolsky, Rosen, 1935] formulated an idea whose essence can be reduced to the following using the example of elementary particles. A quantum object, which can be, for example, two coupled photons, retains some informational connection during the separation (entangle effect). The quantum state of one photon, e.g. its polarization or spin can be immediately transferred to the other photon, which becomes the analog of the first photon, which collapses, disappears, and vice versa. The distance between photons can be any distance. This has been called the Einstein-Podolsky-Rosen (EPR) effect, paradox or channel. Accepted as a synonym for this phenomenon is "quantum nonlocality" (Quantum NonLocality) is also an expression that emphasizes the instantaneous distribution and non-locality in the space of states connected by the quantum states of elementary particles. The principle of causality seems to be violated - effect and cause are not separated by time if time is understood as the organization of a sequence of events. Therefore, Einstein and his co-authors - since they did not have knowledge of the complex structure of time (for example, its fractality) - assessed their purely theoretical, yet rigidly formalized model as not applicable in practice and experiments. This state of contradiction between theory and visible physical reality lasted for about 30 years. Then D. Bell [Bell, 1964; Bell, 1976] developed the idea of EPR at a modern level as well. C. Bennet and his co-authors [Bennet et al, 1993] were also actively involved. The main difficulty was that the basic principle of quantum mechanics derived by Geisenberg regarding the dualistic real-wave state of quantum objects should not be violated in their theoretical constructions. This uncertainty principle states that it is impossible to correctly measure, for example, the properties of a photon as a wave and as an elementary particle at the same time. This problem was eliminated after the possibility of the existence of the "entangled" state of elementary particles was experimentally proven.
Perhaps this "entanglement" is the basic basis for the transfer of genetic (and mental) information between organisms, which can be considered a continuum of elementary particles, and in which the properties of the micro level are reflected on the macro level. In such a detached state, both particles remain part of the same quantum system, so whatever we do to one of them has a predictable effect on the other.
another. Bennett and his colleagues believe that the entangled particles, if they are separated in space, can serve each other as mutual "carriers" of their state and information, since the state of a particle is already information. In this case, however, the information must be understood extremely broadly - like any change. The experimental implementation of the EPR channel required the coexistence of three phonons – one entangled and two plane ones – which was achieved through the work of two research groups – the Viennese group led by Anton Zeulinger and the Roman group led by Francesco De Martini. Experiments by Zeulinger's group [Bouwmeester et al, 1997] demonstrated the practical feasibility of EPR principles for the transfer of polarization states between two photons through a light guide to a third photon over a distance of up to 10 kilometers. Following this discovery, powerful programs are being discussed in leading countries to apply this effect to the development of quantum optical computers, where photons will be the information carriers. Their operating speed and amount of information will be tens of orders of magnitude greater than that of current computers.
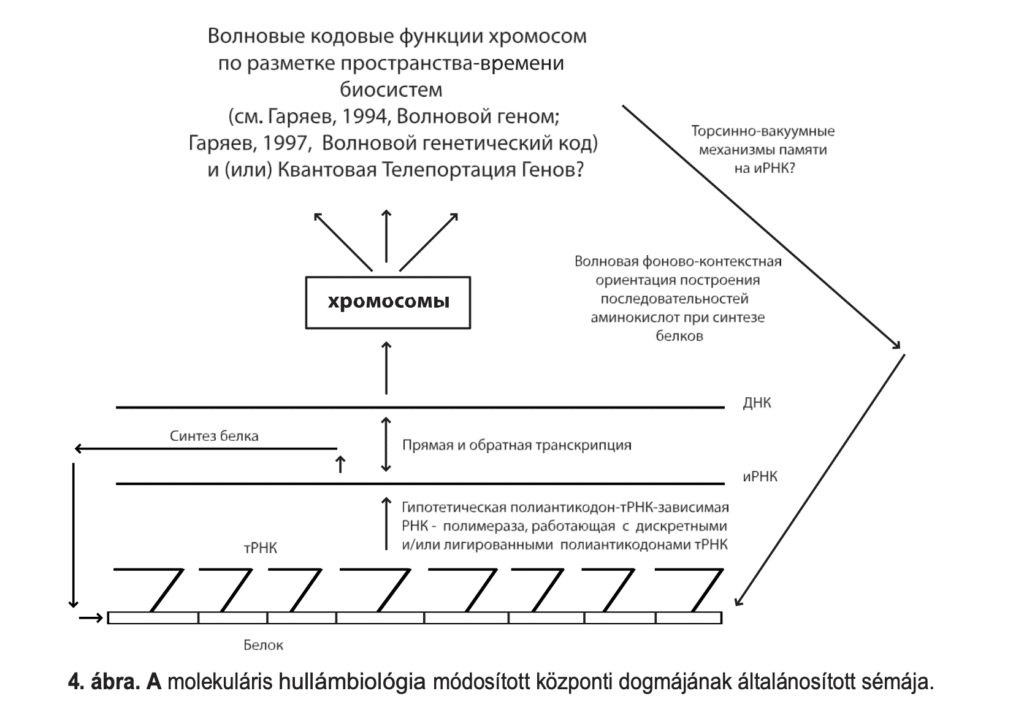
The idea of using the phenomenon of quantum nonlocality by biological systems is very attractive both in perspective and in practice. It corresponds well to our data on the distribution of wave signals in the geno-informational-metabolic and mental areas of biosystems. In this sense, the first, albeit rather weak, attempt to understand the applicability of the EPR concept to biological systems was made somewhat earlier [Josephson, Pallikari-Viras, 1991]. In this work, the theoretical analysis is mainly limited to the statement that the reality perception of living beings is based on a different and in a certain sense more effective principle than that used by the more formal procedures of science. According to the authors, this principle is realized under certain conditions in non-physical, non-static intercommunication signal interactions between distant biosystems, i.e. telepathy. Let's ask the question once more, but in a narrower circle, and without hastily touching on the problem of telepathy - does the phenomenon of quantum nonlocality work in the operation of the genetic apparatus of higher biosystems? If so, in what way? It is clear that here even the assumptions will be of a purely preliminary nature, however, the need for working hypotheses is now due. In wave versions of genome work [Garjajev, 1994, 1997], the EPR effect is a desirable (but not mandatory) link that can logically close the chain of thinking about genome VIM functions. The assumed wave trajectories of chromosome work explain how the spatio-temporal structure of higher biosystems is constructed by the wave and semantic vectors of the work of the heredity apparatus. Such vectors operate through the mechanisms of the holographic memory of the chromosome continuum and the quasi-speech pathways of the construct.
DNA-RNA-Proteins. The genome-biocomputer is read and scanned here, for example, due to the endogenous laser radiation and soliton excitation of the genostructures. The non-locality of the genome, as the chromosomal continuum that encodes and alienates genetic information, is already embedded in its homographic functions. This type of information is distributed throughout the genome as a hologram and/or quasi-hologram and as a fractal at the same time. This can be done if the genome is examined from purely material positions. Quantum wave non-locality does not yet work at this level of genetic information. If the genologram is "read" like a wave, it leads to the fact that the material of the chromosomes alienates signal-shaped wave fronts as the controllers of morphogenesis. This is especially necessary to maintain a stable spatio-temporal structure of the biosystem. For this purpose, the genome generates a kind of "ideal" (wave) model step by step and layer by layer - a plan of the possible material structures of the organism. This is only one of the VIM trends in the construction of the multidimensional structure of the biosystem. In this view, the model of the material-wave organization of biosystems is not yet complete and needs to be developed.
The EPR mechanism, at least at the level of photon-laser and radio wave processes taking place in the chromosomes and proteins of organisms, can represent a significant addition. Such a way of controlling life processes would give cells and tissues fundamentally new opportunities - for example, the possibility of almost instantaneous transfer of huge information sets between all cells and tissues of the biosystem through the polarization channel of photons and radio waves. If this pathway is real, then it becomes clear why biomolecules of strategic importance - nucleic acids and proteins - have the L-isomeric composition of the components, helical twisting and, accordingly, the pronounced dispersion ability of optical rotation, circular dichroism and birefringence . The fact of iso-dimensional quantization of bioorganic molecules is also interpreted differently. The asymmetry of the atoms of bioorganic molecules and the subsequent isomerism is the possibility of rapid self-release of polarization, holographic and other material wave information by the biosystem about the state of its own metabolism and the current space-time structure.
From this point of view, the dual radiation ability of PrPsc aggregates (see above), i.e. the modulation of the polarization vectors of the internal information photon flow through the growing PrPsc protein mass in the brain, which is abnormal from the point of view of the biosystem, acquires an unexpected importance in explaining the mechanisms of prion pathogenesis.
It is typical that the success of experimental quantum teleportation was achieved especially because waveguides (light guides) were used to produce, dilute and “program” photons in space.
UV pumped lasers and polarizers. The bioanalogs of the above components are the microtubules of the cell nucleus and cytoplasm, coherent DNA and chromosomes. The latter are also informational biopolarizers of their own laser radiation, and our direct experiments [Agaltsov, Garyaev et al, 1996] showed that DNA and chromosomes are laser-active media, which was actually confirmed by Japanese researchers, but somewhat differently [Kawabe et al, 2002]. .
Suppose the EPR factor in vivo it functions as a factor controlling the current state of an adult organism from micro and macro level to macro level. But how is it realized in embryogenesis? Perhaps it serves as a mediator of intra- and intra-cellular transfer of DNA-RNA wave copies in different phases of their multifaceted complex operation. It is possible that the VIM effects on DNA preparations independently obtained by us in 1985 and 1991 and by R. Pecora's group in the USA in 1990 are the result of the local quantum teleportation occurring spontaneously during the laser probing of DNA gels due to the dynamic laser light. -scattering spectroscopy. It seems that in this version of the interaction of coherent photons and biostructures, the latter can behave as a liquid crystal system of optically active light guides, which spatially divides polarized photons, with the following exchange of information between them. Another effect is realized in the same system with a new type of memory of genetic structures based on the Fermi-Pasta-Ulam phenomenon. This is accompanied by the appearance of isomorphic temporal autocorrelation functions of light scattering and VIM effects when examining DNA preparations, E. coli 50S subunits and collagen ribosomes [Garyaev, 1994].
If the EPR factor operates in biosystems, it is logical to ask why organisms are not limited to such an efficient form of immediate bioinformational operation. Why would a biosystem need slow nerve impulses? We can only assume that higher organisms needed the nervous system to slow down too fast information processes, which the evolution of the biosphere has not yet achieved. Perhaps the quantum nonlocality functions of the nervous system and the genome complement each other and coexist, sometimes giving rise to human-computer paranormal abilities or telepathy.
Figure 3. Cloning route of in vivo associates (hybrids) of parasite proteins. Proteins 1 + 2 – non-covalent association of subunits, for example in complex proteins such as RNA polymerase.