Le génome multicellulaire est-il quantique non local ?
A.Einstein et ses collègues B.Podolsky et N.Rosen [Einstein, Podolsky, Rosen, 1935] ont formulé une idée dont l'essence peut être réduite à la suivante en utilisant l'exemple des particules élémentaires. Un objet quantique, qui peut être, par exemple, deux photons couplés, conserve une certaine connexion informationnelle pendant la séparation (effet d'enchevêtrement). L'état quantique d'un photon, par ex. sa polarisation ou spin peut être immédiatement transférée à l'autre photon, qui devient l'analogue du premier photon, qui s'effondre, disparaît et vice versa. La distance entre les photons peut être n’importe quelle distance. C'est ce qu'on appelle l'effet, le paradoxe ou le canal Einstein-Podolsky-Rosen (EPR). Accepté comme synonyme de ce phénomène est "non-localité quantique" (Quantum NonLocality) est aussi une expression qui met l'accent sur la distribution instantanée et la non-localité dans l'espace des états reliés par les états quantiques des particules élémentaires. Le principe de causalité semble violé : effet et cause ne sont pas séparés par le temps si le temps est compris comme l'organisation d'une séquence d'événements. Par conséquent, Einstein et ses co-auteurs - puisqu'ils n'avaient aucune connaissance de la structure complexe du temps (par exemple, sa fractalité) - ont évalué leur modèle purement théorique, mais rigidement formalisé, comme non applicable dans la pratique et les expériences. Cet état de contradiction entre la théorie et la réalité physique visible a duré environ 30 ans. Puis D. Bell [Bell, 1964 ; Bell, 1976] a également développé l'idée de la REP à un niveau moderne. C. Bennet et ses co-auteurs [Bennet et al, 1993] ont également été activement impliqués. La principale difficulté était que leurs constructions théoriques ne devaient pas violer le principe de base de la mécanique quantique dérivé de Geisenberg concernant l'état dualiste d'onde réelle des objets quantiques. Ce principe d'incertitude stipule qu'il est impossible de mesurer correctement, par exemple, les propriétés d'un photon en tant qu'onde et en tant que particule élémentaire en même temps. Ce problème a été éliminé après que la possibilité de l'existence de l'état « intriqué » des particules élémentaires ait été prouvée expérimentalement.
Peut-être que cet « enchevêtrement » est la base fondamentale du transfert d’informations génétiques (et mentales) entre organismes, qui peuvent être considérés comme un continuum de particules élémentaires, et dans lequel les propriétés du niveau micro se reflètent au niveau macro. Dans un tel état de détachement, les deux particules font toujours partie du même système quantique, donc tout ce que nous faisons à l’une d’elles a un effet prévisible sur l’autre.
un autre. Bennett et ses collègues pensent que les particules intriquées, si elles sont séparées dans l'espace, peuvent se servir mutuellement de « porteurs » mutuels de leur état et de leurs informations, puisque l'état d'une particule est déjà une information. Dans ce cas, cependant, l’information doit être comprise de manière extrêmement large – comme tout changement. La mise en œuvre expérimentale du canal EPR a nécessité la coexistence de trois phonons – un intriqué et deux plans – qui a été réalisée grâce au travail de deux groupes de recherche – le groupe viennois dirigé par Anton Zeulinger et le groupe romain dirigé par Francesco De Martini. Les expériences du groupe de Zeulinger [Bouwmeester et al, 1997] ont démontré la faisabilité pratique des principes EPR pour le transfert d'états de polarisation entre deux photons via un guide de lumière vers un troisième photon sur une distance allant jusqu'à 10 kilomètres. Suite à cette découverte, de puissants programmes sont en cours de discussion dans les principaux pays pour appliquer cet effet au développement d'ordinateurs optiques quantiques, où les photons seront les supports d'informations. Leur vitesse de fonctionnement et la quantité d'informations seront des dizaines d'ordres de grandeur supérieures à celles des ordinateurs actuels.
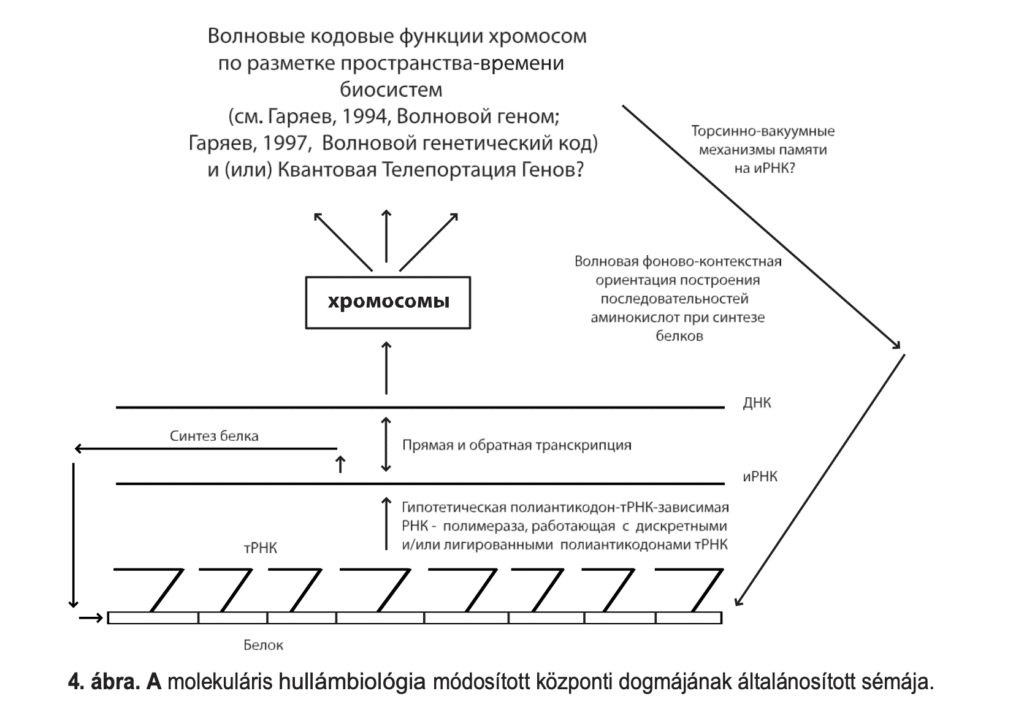
L'idée d'utiliser le phénomène de non-localité quantique par les systèmes biologiques est très attractive tant en perspective qu'en pratique. Cela correspond bien à nos données sur la distribution des signaux d’ondes dans les domaines géno-informationnels-métaboliques et mentaux des biosystèmes. En ce sens, la première tentative, quoique plutôt faible, de comprendre l’applicabilité du concept de REP aux systèmes biologiques a été faite un peu plus tôt [Josephson, Pallikari-Viras, 1991]. Dans ce travail, l'analyse théorique se limite principalement à l'affirmation selon laquelle la perception de la réalité des êtres vivants repose sur un principe différent et, dans un certain sens, plus efficace que celui utilisé par les procédures scientifiques plus formelles. Selon les auteurs, ce principe est réalisé sous certaines conditions dans les interactions de signaux d'intercommunication non physiques et non statiques entre des biosystèmes distants, c'est-à-dire la télépathie. Posons à nouveau la question, mais dans un cercle plus restreint, et sans aborder hâtivement le problème de la télépathie : le phénomène de non-localité quantique agit-il dans le fonctionnement de l'appareil génétique des biosystèmes supérieurs ? Si oui, de quelle manière ? Il est clair qu'ici même les hypothèses seront de nature purement préliminaire, cependant, le besoin d'hypothèses de travail se fait désormais sentir. Dans les versions vagues des travaux sur le génome [Garjajev, 1994, 1997], l’effet EPR est un maillon souhaitable (mais pas obligatoire) qui peut logiquement fermer la chaîne de réflexion sur les fonctions VIM du génome. Les trajectoires ondulatoires supposées du travail chromosomique expliquent comment la structure spatio-temporelle des biosystèmes supérieurs est construite par les vecteurs ondulatoires et sémantiques du travail de l'appareil héréditaire. De tels vecteurs fonctionnent à travers les mécanismes de la mémoire holographique du continuum chromosomique et les voies quasi-verbales de la construction.
ADN-ARN-Protéines. Le génome-bioordinateur est lu et scanné ici, par exemple, grâce au rayonnement laser endogène et à l'excitation des solitons des génostructures. La non-localité du génome, en tant que continuum chromosomique qui code et aliène l’information génétique, est déjà ancrée dans ses fonctions homographiques. Ce type d'information est distribué dans tout le génome sous forme d'hologramme et/ou quasi-hologramme et de fractale à la fois. Cela est possible si le génome est examiné à partir de positions purement matérielles. La non-localité des ondes quantiques ne fonctionne pas encore à ce niveau d’information génétique. Si le génologramme est « lu » comme une onde, cela conduit au fait que le matériau des chromosomes aliène les fronts d'onde en forme de signal en tant que contrôleurs de la morphogenèse. Ceci est particulièrement nécessaire pour maintenir une structure spatio-temporelle stable du biosystème. À cette fin, le génome génère une sorte de modèle « idéal » (onde) étape par étape et couche par couche - un plan des structures matérielles possibles de l'organisme. Ce n’est qu’une des tendances VIM dans la construction de la structure multidimensionnelle du biosystème. De ce point de vue, le modèle de l’organisation des ondes matérielles des biosystèmes n’est pas encore complet et doit être développé.
Le mécanisme EPR, au moins au niveau des processus photons-laser et ondes radio se déroulant dans les chromosomes et les protéines des organismes, peut représenter un ajout significatif. Une telle manière de gérer les processus vitaux offrirait aux cellules et aux tissus des opportunités fondamentalement nouvelles - par exemple, la possibilité de transfert presque instantané d'énormes ensembles d'informations entre toutes les cellules et tous les tissus du biosystème via le canal de polarisation des photons et des ondes radio. Si cette voie est réelle, il devient alors clair pourquoi les biomolécules d'importance stratégique - acides nucléiques et protéines - ont une composition en isomère L des composants, une torsion hélicoïdale et, par conséquent, une capacité de dispersion prononcée de rotation optique, de dichroïsme circulaire et de biréfringence. Le fait de la quantification isodimensionnelle des molécules bioorganiques est également interprété différemment. L'asymétrie des atomes des molécules bioorganiques et l'isomérie qui en résulte sont la possibilité d'une auto-libération rapide d'informations de polarisation, holographiques et d'autres ondes matérielles par le biosystème sur l'état de son propre métabolisme et la structure spatio-temporelle actuelle.
De ce point de vue, la double capacité de rayonnement des agrégats de PrPsc (voir ci-dessus), c'est-à-dire la modulation des vecteurs de polarisation du flux de photons d'information interne à travers la masse croissante de protéines PrPsc dans le cerveau, ce qui est anormal du point de vue de le biosystème, acquiert une importance inattendue dans l'explication des mécanismes de la pathogenèse des prions.
Il est typique que le succès de la téléportation quantique expérimentale ait été obtenu notamment grâce à l'utilisation de guides d'ondes (guides optiques) pour produire, diluer et « programmer » des photons dans l'espace.
Lasers et polariseurs pompés UV. Les bioanalogues des composants ci-dessus sont les microtubules du noyau et du cytoplasme cellulaire, l'ADN cohérent et les chromosomes. Ces derniers sont également des biopolariseurs informationnels de leur propre rayonnement laser, et nos expériences directes [Agaltsov, Garyaev et al, 1996] ont montré que l'ADN et les chromosomes sont des milieux actifs au laser, ce qui a été confirmé par des chercheurs japonais, mais de manière quelque peu différente [Kawabe et al, 2002]. .
Supposons que le facteur EPR in vivo il fonctionne comme un facteur contrôlant l’état actuel d’un organisme adulte du niveau micro et macro au niveau macro. Mais comment se réalise-t-elle dans l’embryogenèse ? Peut-être sert-il de médiateur du transfert intra- et intra-cellulaire des copies d'ondes ADN-ARN dans différentes phases de leur fonctionnement complexe à multiples facettes. Il est possible que les effets VIM sur les préparations d'ADN obtenus indépendamment par nous en 1985 et 1991 et par le groupe de R. Pecora aux États-Unis en 1990 soient le résultat de la téléportation quantique locale se produisant spontanément lors du sondage laser des gels d'ADN en raison de la dynamique lumière laser.-spectroscopie de diffusion. Il semble que dans cette version de l'interaction de photons cohérents et de biostructures, ces dernières puissent se comporter comme un système à cristaux liquides de guides de lumière optiquement actifs, qui divise spatialement les photons polarisés, avec l'échange d'informations qui s'ensuit entre eux. Un autre effet est réalisé dans le même système avec un nouveau type de mémoire de structures génétiques basé sur le phénomène Fermi-Pasta-Ulam. Ceci s'accompagne de l'apparition de fonctions d'autocorrélation temporelle isomorphes de diffusion de la lumière et d'effets VIM lors de l'examen de préparations d'ADN, des sous-unités 50S d'E. coli et des ribosomes de collagène [Garyaev, 1994].
Si le facteur EPR opère dans les biosystèmes, il est logique de se demander pourquoi les organismes ne se limitent pas à une forme aussi efficace d’opération bioinformationnelle immédiate. Pourquoi un biosystème aurait-il besoin d’influx nerveux lents ? Nous ne pouvons que supposer que les organismes supérieurs avaient besoin du système nerveux pour ralentir les processus d'information trop rapides, ce que l'évolution de la biosphère n'a pas encore réalisé. Peut-être que les fonctions de non-localité quantique du système nerveux et du génome se complètent et coexistent, donnant parfois naissance à des capacités paranormales ou à la télépathie entre humains et ordinateurs.
Figure 3. Voie de clonage d'associés in vivo (hybrides) de protéines parasitaires. Protéines 1 + 2 – association non covalente de sous-unités, par exemple dans des protéines complexes telles que l'ARN polymérase.